中华白海豚(Sousa chinensis)为沿岸河口定栖性小型齿鲸类, 又名印度太平洋驼背豚(Indo-Pacific humpback dolphin), 属海豚科(Delphinidae)白海豚属(Sousa), 1988年被国务院列为国家一级保护动物[1]。2008年国际自然及自然保护联盟(IUCN)将中华白海豚定为近危等级物种[2], 同时, 其也被濒危野生动植物国际贸易公约(CITES)附录Ⅰ收录[3], 属于禁止贸易物种。
中华白海豚主要分布于西太平洋和印度洋的沿岸水域[4], 在中国水域主要分布于东南沿海的河口区[5]。据公开的报道, 中国沿海有中华白海豚分布的水域为:珠江口[6-12]、厦门[13-17]、湛江东部海域[18]、北部湾北部沿岸[19-22]、台湾西南部沿岸[23-24]、潮州—汕头[25], 以及李松海等人在海南三亚西南部也发现了中华白海豚的分布[26]。上述报道对中国沿海中华白海豚的研究主要侧重于分布、数量、个体识别及家域等种群基础生物学方面, 有关栖息地选择, 特别是环境因子等如何影响中华白海豚的活动或迁移等的研究还比较缺乏, 仅见个别报道[15, 27], 而栖息地选择的研究对中国沿海中华白海豚的保护规划和管理有很大的助益。珠江口中华白海豚种群的分布范围包括了珠江东部入海口(即伶仃洋)和西部入海口(西部河口区)冲淡水影响的区域, 从伶仃洋东部的深圳、香港水域向西延伸至江门市的上、下川岛水域, 分布范围超过3000km2, 种群规模超过2500头, 被认为是目前全球范围内所知最大的种群[11]。其栖息的珠江河口周边沿岸城市密集, 密布着广州、深圳、香港、珠海和澳门等城市, 这些城市经济发达、人口密集, 频繁的人类活动(如航运、填海造地等)加剧了中华白海豚栖息地环境的变化, 使其生存环境变得更加复杂。研究该种群的分布或活动与环境因子的关系, 可以有效的了解中华白海豚对栖息地条件的要求, 预测其分布范围和潜在的重要栖息地, 为中华白海豚的保护管理提供依据, 如减少潜在栖息地的破坏及人类活动的干扰、栖息地的修复等。同时, 珠江西部河口属开放型的河口区, 有别于属于半封闭型海湾的伶仃洋区域, 这与中国沿海甚至世界其他种群的栖息地有很大的相似性, 以珠江西部河口区作为研究区域也具有一定的代表性。
海洋鲸豚类动物的活动范围较大, 由于活动能力较强而难以追踪, 为研究其季节迁移带来一定的困难。研究表明, 在不同研究尺度下, 海洋鲸豚类动物的分布与环境要素相关, 这些要素包括自然地理要素(水深等)和海洋要素(水温和叶绿素浓度等)[28-29]。这些因素对海豚的影响既有直接的, 又有间接的。水温对鲸豚类分布既有直接影响也有间接影响, 直接影响主要体现在自身体温调节的能量成本[30], 间接影响表现为对鱼类、头足类和浮游动物等饵料的分布。而鱼类和头足类的分布则与海洋水文特征及环境要素相关, 包括水深、上升流和海洋锋面等。鲸豚类的饵料生物在不同时间有水平移动也有垂直移动, 这使得在小尺度时空范围内很难预测其栖息地使用模式。在小尺度范围内, 测定环境参数要比测定饵料生物更为方便。环境参数可以较好的模拟小尺度范围内鲸豚类的栖息地偏好[31]。了解中华白海豚的分布与环境因子的关系, 可以识别其栖息地使用要求和分布模式, 提供更详细的捕食习性和识别潜在的重要栖息地。
GAM(Generalized Additive Models, 广义加性模型)模型首先由Hastie等[32]提出, 是一种非线性关系。GAM模型分析方法已用来分析海洋经济鱼种分布与地理及环境变量的关系[33-36], 应用于海洋鲸豚类栖息地选择分析也越来越普遍[37-38]。在珠江西部河口水域, 已有关于中华白海豚的分布及其季节迁移趋势的报道[12], 但目前还没有针对珠江西部河口中华白海豚的分布与海洋环境因子(包括其他游泳生物等)关系的研究。本文使用GIS和GAM统计模型, 利用2012年珠江口西部河口的海豚截线观测、渔业资源及环境要素定点采样数据, 分析包括以下3个方面的内容:(1)珠江口西部河口渔业资源的密度分布;(2)中华白海豚分布与环境因子的关系(水深、底层水温、盐度、pH值、溶解氧和离岸距离等);(3)中华白海豚分布与其他游泳生物资源的关系。
1 材料与方法 1.1 调查范围本次调查分为海豚截线观测和渔业资源及环境要素定点采样调查, 观测截线和渔业资源及环境要素采样站位见图 1。调查海区位于珠江口西部河口区域, 东起珠海市横琴岛西南, 西至江门市下川岛以东, 覆盖磨刀门、鸡啼门、虎跳门和崖门等珠江西部四大入海口门。调查区域设置了一系列的平行观测截线, 截线由岸边水深3m附近(调查船吃水深度2.5m, 略大于2.5m以保证航行安全)向南延伸至20m等深线附近, 截线间隔约4km, 预设截线总长约440km, 调查范围为:112.67°—113.5°E、21.5°—22°N。
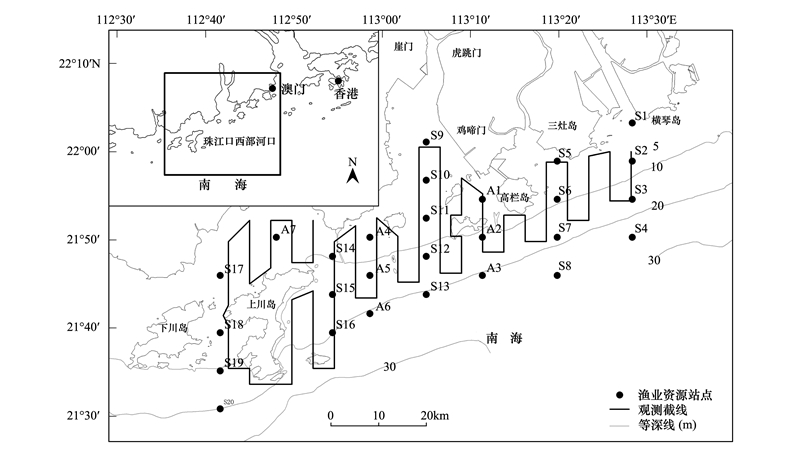 图 1 2012年珠江口西部河口海豚观测截线和渔业资源、环境要素采样站位
Fig. 1 Transect lines of dolphin survey, as well as with fisheries and environmental factors sampling stations in the western Pearl River Estuary during 2012
S:渔业资源和环境要素采样站点;A:渔业资源和环境要素采样站点
图选项
1.2 调查方法
图 1 2012年珠江口西部河口海豚观测截线和渔业资源、环境要素采样站位
Fig. 1 Transect lines of dolphin survey, as well as with fisheries and environmental factors sampling stations in the western Pearl River Estuary during 2012
S:渔业资源和环境要素采样站点;A:渔业资源和环境要素采样站点
图选项
1.2 调查方法
本次调查分为海豚调查和渔业资源及环境要素调查, 时间为2012年2—12月, 于丰水期(5月和8月)和枯水期(2月和12月)各进行了2个航次, 即海豚和渔业资源等均进行了4个航次的同步调查。中华白海豚观测以船基截线法进行, 目视观测和数据记录依据Jefferson等[6-7]方法进行。使用粤东莞00589渔船作为观测船, 观测台设于船艏上甲板, 目视点离水面约4—5m。在适宜的观测条件下(蒲福海况0—5级, 无大雨, 能见度不小于1200m), 船只以13—15km/h的航速沿预设截线航行, 由2人组成一班同时观测, 主观察员用内置指南针双目望远镜(Nikon7×50 IF WP)观察, 副观察员用肉眼观察兼数据记录, 每隔0.5h按顺序轮换主观察员。观测记录包括截线观测记录和海豚目击记录:截线观测记录主要包括观测开始和结束的时间和位置、航速、航向、海况、能见度和航程等, 发现海豚将结束一个观测系列, 同时估算海豚个体数、群体组成等;海豚目击记录包括首次目击的时间、位置、角度、目测距离、个体数、组成和行为等。位置、航速和航程均由GPS(Magellan explorist 210)获得, 航向和角度由望远镜测得。
渔业资源及环境要素采样调查与海豚截线调查同步进行。在海豚观测截线范围内设置7个剖面由浅水区至深水区纵向分布的系列采样站点, 共27个, 分列于不同的水深梯度(5—20m)。使用底层单拖渔船, 采用定点拖网的方式, 在海豚调查的同一天进行拖网采样。每个站位拖网一次, 船速为3—4节, 每网拖30min, 拖网距离约3km。起网后, 在现场对渔获物全部取样分析, 包括种类鉴定和生物学测定。种类分类鉴定到种, 每个种类均测定样品的总重量、总尾数以及最小、最大体长和最小、最大体重。在拖网前进行环境要素的采样, 包括底层水温、盐度、pH、DO(溶解氧, Dissolved oxygen), 渔业环境要素采用YSI 556 MPS型多功能水质仪在现场测定。
1.3 数据处理和分析将研究区域沿着观测截线分为4km×8km网格, 这样折中的处理可以减少大多数网格没有海豚观测数据, 同时又可与环境要素建立联系, 基于网格的研究方法可以减少数据上的自动相关性这个潜在问题。研究区域分为56个网格, 覆盖区域为112.67°—113.5°E、21.5°—22°N。由北向南将所有网格用不同颜色程度分为5个子区域, 依次为SA1, SA2, SA3, SA4和SA5(区域网格见图 2)。为了便于海豚观测数据的定量分析, 因海豚观测首先以群体作为发现目标, 引入群体观测努力量SPUE(sightings per 100km search effort), 即每100km的截线观测努力目击到的海豚群数, 简称海豚目击率。利用GAM模型分析海豚目击率与环境因子关系。根据调查经验, 选取9个可利用的环境数据及可能影响海豚目击率的因子:海况、水深、底层水温、盐度、pH值、溶解氧、捕食种密度、游泳生物密度和离岸距离。香港水域的研究表明, 石首鱼科、带鱼科、鳀科和鲱科鱼类占其捕食消耗的93%[39]。在这些科的鱼类当中, 叫姑鱼(Johnius grypotus)、棘头梅童鱼(Collichthys lucidus)、汉氏棱鳀(Thrissa hamiltonii)等河口鱼类是中华白海豚最为喜好的捕食鱼类, 本文所定义的捕食种密度指的是上述鱼类的分布情况。
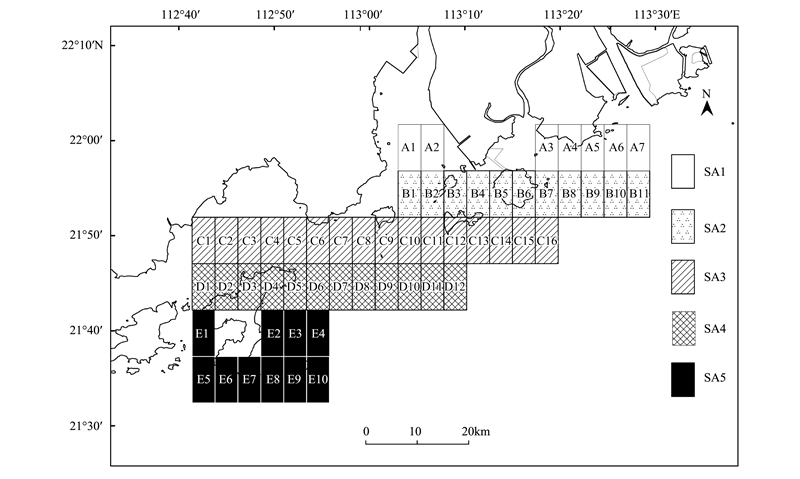 图 2 调查区域网格图
Fig. 2 Map of grids on survey waters
A、B、C、D、E:网格系统The grid system; SA:网格系统子区域Sub-areas of the grid system; 自北向南依次是SA1,SA2,SA3,SA4和SA5
图选项
图 2 调查区域网格图
Fig. 2 Map of grids on survey waters
A、B、C、D、E:网格系统The grid system; SA:网格系统子区域Sub-areas of the grid system; 自北向南依次是SA1,SA2,SA3,SA4和SA5
图选项
环境因子之间可能存在共线性关系, 分析数据时, 采用Pearson相关系数判定两者之间的相关性, 当两者存在共线性关系时, 只选取其中一个变量为解释变量[40]。
采用扫海面积法对研究区域渔业资源进行资源量评估[41], 计算公式如公式(1):
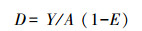 (1)
(1) 式中, D为标准资源量(kg/km2)、A为每小时扫海面积(km2/h)、Y为平均捕捞率(kg/h)、E为逃跑率, 本研究E取值为0.5。
1.4 广义加性模型(GAM)GAM模型是广义线性模型(GLM, Generalized linear models)的推广, 方程的一般形式为公式(2):
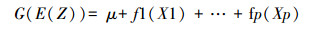 (2)
(2) 式中, G连接函数, E为期望, Z为应变量, X为自变量, f为平滑函数(如样条平滑函数、局部平滑函数), μ截距, p为参数个数。
本研究分别利用GAM模型建立调查期间每个网格的海豚目击率与环境因子的关系;每一次海豚目击率变化与环境因子的关系。利用逐步回归方法(R中的mgcv软件包[42]), 采用AIC(Akaike Information Criterion)信息准则和各解释变量的显著程度(P值)对GAM模型进行选择[43]。基于对数据的分析, 本研究采用Possion分布, 自然对数为连接函数。
2 结果 2.1 渔业资源调查结果采用扫海面积法估算研究区域游泳动物的资源密度。根据公式计算, 该海域枯水期游泳动物的资源密度介于667.88—2679.94kg/km2之间, 平均1444.41kg/km2, 各站资源密度见表 1, 丰水期游泳动物的资源密度介于227.03—4085.22kg/km2之间, 平均1573.57kg/km2, 各站资源密度见表 1。本次调查共记录游泳动物200种, 隶属19目81科。其中鱼类15目61科137种;甲壳类2目16科55种;头足类2目4科8种。
表 1 调查区域枯水期和丰水期游泳生物密度/(kg/km2) Table 1 Stock density of nektons in survey waters in dry season and wet season 站位Station 枯水期
Dry season 比例
Percent/% 丰水期
Wet season 比例
Percent/% A2 1959.83 17.77 871.00 1.18 A3 2083.86 4.18 1187.63 10.99 A4 1479.07 13.49 2365.61 13.86 A5 1930.80 13.69 2442.81 9.26 A6 1533.82 12.74 1109.31 22.12 A7 2366.57 6.17 1070.20 11.12 S1 710.54 12.20 590.82 16.38 S10 667.88 12.91 3896.50 25.46 S11 1675.06 24.09 3343.86 22.48 S12 1607.71 33.32 904.30 7.98 S13 1658.50 63.33 227.03 53.72 S14 937.87 22.76 1002.29 4.49 S15 978.54 20.39 782.99 13.04 S16 1274.74 26.59 730.40 12.19 S17 2223.97 12.82 4085.22 17.00 S18 854.55 29.31 1337.39 43.61 S19 1196.99 13.84 944.66 20.13 S2 2148.33 19.52 514.52 29.68 S20 1135.29 17.71 1383.96 23.00 S3 1130.89 28.36 3704.49 19.19 S4 682.72 22.91 886.94 28.73 S5 2679.24 9.92 991.24 1.90 S6 1070.11 46.90 883.69 5.99 S7 1494.73 44.09 2955.02 6.39 S8 708.96 44.09 915.43 7.73 S9 1364.00 2.83 1785.49 29.77 表选项 2.2 中华白海豚目击率与环境因子的关系
本研究共获取9个解释变量, 包括海况、水深、底层水温、盐度、pH值、溶解氧、捕食种密度、游泳生物密度和离岸距离。采用Pearson相关系数判定两者之间的相关性, 当两者存在共线性关系时, 只选取其中一个变量为解释变量。捕食种密度、盐度和pH值间的相关系数大于0.5, 因捕食种是海豚的直接食物来源, 对海豚目击率影响可能更明显, 只选取捕食种密度分析;底层水温和溶解氧的相关系数大于0.5, 底层水温比溶解氧受外界影响波动更小, 选取保留的变量为底层水温。最终模型选取6个变量包括水深、离岸距离、海况、底层水温、捕食种密度和游泳生物密度。海豚目击记录位置水深变化在20m以浅水域, 底层水温变化范围为15.75—30.9℃, 盐度变化范围为15.6—33.6, pH值变化范围为6.93—8.15, 溶解氧变化范围为3.3—13.45。海豚目击率最大值的水深变化范围为5—10m。
GAM模型中各个因子的显著性和AIC值见表 2, 结果显示, 选取的6个解释变量均极显著(P值均小于0.05)。图 3反映了海豚目击率与水深之间的关系, 水深范围在5—10m变化时, 随着水深增加, 海豚目击率单调递增, 当水深范围在10—15m变化时, 海豚目击率变化较小, 但当水深大于15m后, 海豚目击率单调递减; 反映了海豚目击率与离岸距离的关系, 当离岸距离范围在0.1—3km时, 随着离岸距离增加, 海豚目击密度单调递增;当离岸距离大于3km时, 海豚目击率单调递减; 反映了海豚目击率与底层水温之间的关系, 当底层水温范围在17—19.5℃时, 曲线波动变化较小;当底层水温大于19.5℃时, 曲线递减明显, 分析数据也发现, 此处方格主要位于海豚调查截线的边缘, 也即处在20m等深线附近; 反映了海豚目击率与海况的关系, 随着海况增加, 海豚目击率单调递减, 近似为直线。
表 2 GAM模型拟合结果的偏差分析 Table 2 Analysis of deviance for generalized additive models (GAMs) fitted to the dolphin encounter rate data 模型因子Model factors 残差自由度
Residual degree of freedom 估计自由度
Estimated degree of freedom AIC值
AIC value 累计解释偏差
Cumulation of deviance explained/% F检验
Pr(F) 偏差
Deviance 游泳生物密度
Density of CWD′s prey fish/(kg/km2) 49.06 2.939 381.62 19.7 8.3×10-15*** 307.25 水深Depth/m 47.04 1.977 296.73 43 1.75×10-12*** 217.97 捕食种密度
Density of CWD′s prey fish/(kg/km2) 46.09 2.7087 285.81 50.6 0.000311*** 205.49 离岸距离Distance to land/km 43.54 1.61 273.39 56 0.000335*** 187.88 底层水温Temperature/℃ 40.15 3.0 259.29 61.4 0.00511** 167.1 海况Beaufort sea state 38.27 1.011 242.36 64.7 0.00109** 146.41 Pr(F)指对本行模型与上一行模型进行离差分析时F检验所获得的P值:*P < 0.05, **P < 0.01, ***P < 0.001;CWD:中华白海豚China white dolphin 表选项
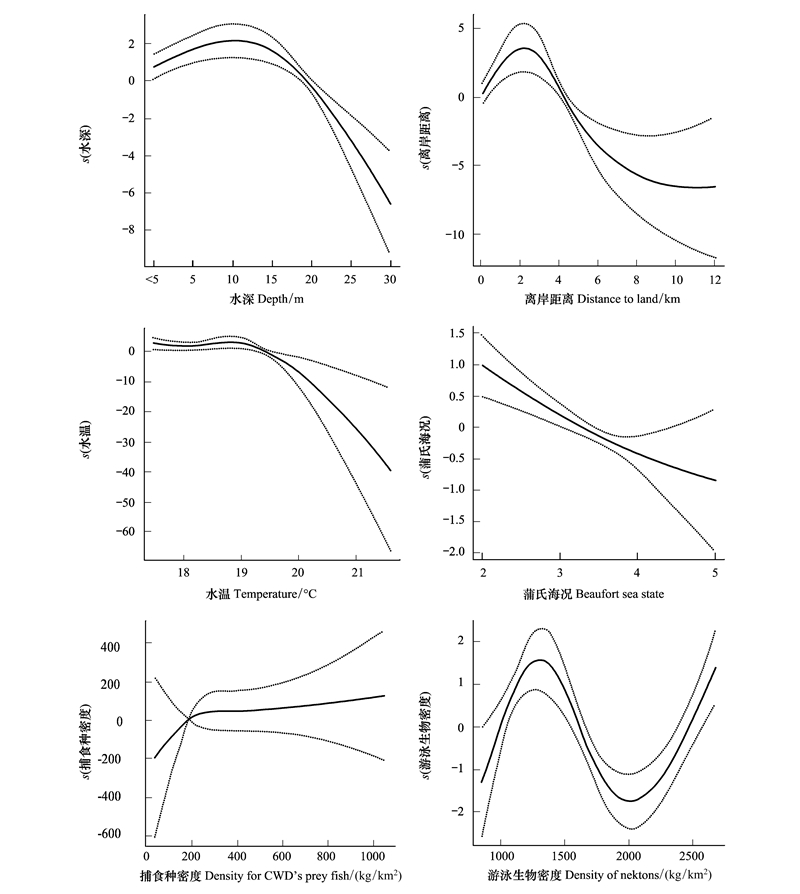 图 3 中华白海豚群体目击率与环境因子的GAM分析
Fig. 3 The relationships between the encounter rate of humpback dolphin and its related environmental factors from GAM
图选项
图 3 中华白海豚群体目击率与环境因子的GAM分析
Fig. 3 The relationships between the encounter rate of humpback dolphin and its related environmental factors from GAM
图选项
图 3可见, 海豚目击率与游泳生物密度之间存在非线性关系, 当游泳生物密度为850—1250kg/km2范围时, 随着游泳生物密度增加, 海豚目击率单调递增;当游泳生物密度为1250—2000kg/km2范围时, 随着游泳生物密度增加, 海豚目击率单调递减;当游泳生物密度大于2000kg/km2范围时, 随着游泳生物密度增加, 海豚目击率单调递增; 反映了海豚目击率与捕食种密度的关系, 当捕食种密度范围在38—250kg/km2时, 随着捕食种密度的增加, 海豚目击率单调递增;当捕食种密度大于250kg/km2时, 曲线趋于平缓, 变化较小。
3 讨论广义加性模型(GAM)无需限定变量的潜在关系, 提供了灵活的、非线性框架来研究种群—环境模型。本研究中多数潜在关系是非线性的, 表明GAM模型是适合的。对于珠江口西部河口中华白海豚GAM模型, 利用海豚目击率为环境因子的函数, 某种程度上有说服力。
海洋鲸豚类动物对栖息地及生存环境变化有选择性[44], 在高生产力区、中尺度海洋锋面和上升流地区, 中上层肉食动物变动与环境因素均已有研究, 鲸豚类数量变动与叶绿素浓度相关[45-46]。研究鲸豚类数量变动时空关系与环境因子一般通过间接方法, 不同的研究尺度结果差异较大。鲸豚类生物通常以鱼类等游泳生物为食, 充分了解研究区域的水文要素和海流系统、以及鲸豚类的习性易于发现数量变动关系。
3.1 海豚目击率与其他游泳生物的关系本研究通过渔业资源调查, 共记录游泳生物200种, 隶属19目81科。其中鱼类15目61科137种;甲壳类2目16科55种;头足类2目4科8种。获得海豚主要食物鱼类共26种, 分别有花鰶(Clupanodon thrissa)、斑鰶(Clupanodon punctatus)、鳓(Ilisha elongata)、印度鳓(Ilisha indica)、金色小沙丁鱼(Sardinella aurita)、雷氏小沙丁鱼(Sardinella richardsoni)、黄鲫(Setipinna taty)、凤鲚(Coilia mystus)、康氏小公鱼(Stolephorus commersoni)、汉氏棱鳀(Thrissa hamiltonii)、长颌棱鳀(Thrissa setirostris)、杜氏棱鳀(Thrissa dussumieri)、皮氏叫姑鱼(Johnius belengeri)、杜氏叫姑鱼(Johnius dussumieri)、白姑鱼(Argyrosomus argentatus)、棘头梅童鱼(Collichthys lucidus)、截尾白姑鱼(Argyrosomus aneus)、勒氏短须石首鱼(Umbrina russelli)、银牙 (Otolithes argenteus)、尖头黄鳍牙(Chrysochir aureus)、大黄鱼(Pseudosciaena crocea)、短带鱼(Trichiurus brevis)、带鱼(Trichiurus haumela)和南海带鱼(Trichiurus nanhaiensis)等。不同季节海豚食物鱼类种类和数目有差别。
(Otolithes argenteus)、尖头黄鳍牙(Chrysochir aureus)、大黄鱼(Pseudosciaena crocea)、短带鱼(Trichiurus brevis)、带鱼(Trichiurus haumela)和南海带鱼(Trichiurus nanhaiensis)等。不同季节海豚食物鱼类种类和数目有差别。
本文分析结果中, 海豚目击率与游泳生物密度之间存在非线性关系, 当游泳生物密度为850—1250kg/km2范围时, 随着游泳生物密度增加, 海豚目击率单调递增;当游泳生物密度为1250—2000kg/km2范围时, 随着游泳生物密度增加, 海豚目击率单调递减;当游泳生物密度大于2000kg/km2范围时, 随着游泳生物密度增加, 海豚目击率又单调递增。一般来说海豚目击率多数与游泳生物密度呈正相关, 但分析中却呈现先递增后递减再递增的波动变化, 原因可能与游泳生物渔获中的海豚捕食种所占比例有关, 本调查中海豚捕食种在游泳生物中的比例变化范围为1.9%—63.3%, 即捕食种密度与游泳生物密度并非呈现正相关, 这可能是导致海豚目击率波动的因素。
为消除游泳生物中非中华白海豚捕食种的干扰, 我们进一步单独考察了海豚目击率与捕食种密度的关系。当捕食种密度范围在38—250kg/km2时, 随着捕食种密度的增加, 海豚目击率单调递增;但当捕食种密度大于250kg/km2时, 曲线趋于平缓, 变化较小, 模拟结果说明中华白海豚的活动与捕食种密度密切相关。但是, 模拟结果也提供了另外一个信息, 即当食物来源不多时, 海豚需要花费较多时间在食物较多的水域觅食, 此时捕食种密度对海豚目击率的正面影响较大, 而当水域的食物来源充足时, 海豚只需花费较少的时间就能获得足够的食物, 因此在某一特定水域的逗留时间变短, 此时食物密度因子对海豚目击率的正面影响有所弱化。
3.2 海豚目击率与水深的关系水深对海豚目击率有影响, 水深范围在5—10m变化时, 随着水深增加, 海豚目击率单调递增, 当水深范围时在10—15m变化时, 海豚目击率略呈递减趋势但变化较小, 但当水深大于15m后, 海豚目击率单调递减。水深被认为是限制中华白海豚的离岸分布的要素之一, 在南非水域25m等深线被认定为中华白海豚离岸分布的临界深度[47], 陈涛等于2007—2008年的调查认为20m等深线可能是珠江西部河口中华白海豚离岸分布的边界, 5—10m为其主要分布区, 丰水期趋向5m以浅的近岸水域活动, 而枯水期时趋向10m以深的离岸水域活动[12]。本文的模拟结果与陈涛等数年前的统计分析结果基本一致, 也进一步说明在这一区域, 不同调查年份海豚分布对水深选择的变化不大, 其较为喜欢的水深在10m附近。
3.3 海豚目击率与离岸距离的关系影响海豚目击率因子包括离岸距离, 当离岸距离范围在0.1—3km时, 随着离岸距离增加, 海豚目击密度单调递增;当离岸距离大于3km时, 海豚目击率单调递减。说明珠江口的中华白海豚主要活动于离岸距离小于3km的近岸水域, 而离岸3km以内的近岸水域是人类活动干扰最为剧烈的水域, 如填海造地等活动。Karczmarsk等在非洲Algoa港水域调查表明[47], 海豚分布在距离岸边200—400m, 占目击次数的58.6%, Parra等在澳大利亚昆士兰东北部调查表明[48], 海豚的目击分布离岸距离多小于5km。可能是不同水域地形环境的离岸活动范围会有所差异, 珠江西部河口中华白海豚离岸分布的临界深度要小于南非水域, 但其离岸分布距离要远大于南非水域, 而小于澳大利亚昆士兰东北沿岸海域。
3.4 海豚目击率与水温的关系水温对于中华白海豚栖息地偏好及分布影响已有研究, 但选取的水层多为表层水温, 结果表明表层水温对其栖息地偏好和分布无显著影响[15, 27]。本文的研究选择了底层水温, 底层水温受气温变化的影响较小, 其波动变化幅度也较表层小。珠江口属典型的河口湾, 夏季底层水温最高为29.2℃, 最低为21.9℃;冬季水温分布比夏季均匀, 温差在2℃左右, 水温变化范围16.6—18.7℃, 表底层水温分布趋势基本相同, 在河口湾附近海域, 水温由岸往外呈递增的变化趋势[49]。在本文的研究中, 当底层水温范围在17—19.5℃时, 海豚目击率的曲线波动变化较小;当底层水温大于19.5℃时, 曲线递减明显。海豚为哺乳动物, 水温变化可能对其直接影响不大, 而鱼类等对水温的变化较为敏感, 因此底层水温变化会直接影响到捕食种鱼类分布的变化, 进而影响到海豚分布。
比较底层水温的空间分布时, 发现本次调查中底层水温大于19.5℃时, 一般位于20m等深线附近。陈涛等曾认为20m等深线可能是珠江西部河口中华白海豚离岸分布的边界[12], 本文的底层水温模拟结果进一步解释了为何是20m等深线, 底层水温可能是主要的限制因子, 底层水温直接限制着捕食种鱼类的分布, 进而影响到海豚的离岸分布。在其他区域, 如珠江东部河口的伶仃洋, 大屿山与桂山岛之间的大濠水道水深超过20m, 但也是中华白海豚使用率较高的区域之一[6], 该水道的底层水温可能不高, 适合中华白海豚捕食种鱼类的栖息。这一现象对判断中国沿海其他区域中华白海豚的离岸分布边界有参考意义。
3.5 海豚目击率与海况的关系海况与海豚群体目击率的关系表现为, 随着海况增加, 海豚群体目击率单调递减, 近似为直线。Jefferson等在香港水域的统计表明[6], 海豚群体的目击率随着海况的增大显著递减, 本文的GAM模拟结果进一步印证了这个统计结果。但是Jefferson的分析也表明, 采用DISTANCE评估海豚的数量时, 当海况为0—3级时评估结果变化不大, 在无更多观测数据的情况下误差仍处于可接受的范围, 但当海况大于3级时评估结果将显著减少, 因此在评估海豚数量时一般选用0—3级的观测数据, 以避免评估结果的偏低。因为当4级及以上蒲福海况时, 海面的白浪较多, 会干扰观测效果, 影响到海豚的目击率。
3.6 保护建议从本文的研究结果可知, 珠江西部河口中华白海豚主要使用离沿岸3km范围以内的水域, 而该水域范围是流刺网、底拖网等捕捞作业的主要水域, 也是沿岸填海主要影响区域。因此, 沿岸的捕捞作业, 以及填海造地对中华白海豚种群的影响巨大。不但会造成重要栖息地的丧失以及食物资源的紧张, 渔业捕捞还会误捕或误伤中华白海豚, 如渔网割伤或缠绕, 加剧种群的生存压力。为了有效地保护这一河口生态系统的旗舰物种, 河口沿岸的填海需谨慎, 限制流刺网等渔业生产渔船数量, 同时对已填海破坏的区域进行生态修复, 通过适宜的人工鱼礁投放和增殖放流等措施保证海豚的食物资源。
标签组:[时政]
上一篇:海豚作文550字